Biosynthesis and signaling of ethylene and their regulation on seed germination and dormancy
SONG Song-Quan,1,3,*, LIU Jun2, XU Heng-Heng2, ZHANG Qi2, HUANG Hui3, WU Xian-Jin31 Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2 Agro-Biological Gene Research Center, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, Guangdong, China 3 Key Laboratory of Research and Utilization of Ethnomedicinal Plant Resources of Hunan Province / College of Biological and Food Engineering, Huaihua University, Huaihua 418008, Hunan, China
This study was supported by the National Science and Technology Support Program.2012BAC01B05 the National Natural Science Foundation of China.31371715 the National Natural Science Foundation of China.31640059 the Guangdong Science and Technology Program.2016B030303007
Abstract Seed germination, a key ecological and agronomic trait, is determined by both internal and external cues that regulate the dormancy status and the potential for germination in seeds, and plays a critical role during the subsequent growth, development and production of plants. Dormancy is the temporary failure of seed germination under favorable conditions. Ethylene is a simple gaseous phytohormone with multiple roles in regulation of metabolism at molecular, cellular, and whole plant levels. It influences performance of plants under optimal and stressful environments by interacting with other signaling molecules. In the present paper, we mainly summarize ethylene biosynthesis and signaling, the role of ethylene in seed germination and dormancy release, and the interaction of ethylene with phytohormone abscisic acid and gibberellin, and propose some scientific problems to be required to investigate further in order to provide an idea for explaining the molecular mechanism of seed germination and dormancy regulated by ethylene. Keywords:abscisic acid;biosynthesis and signaling;crosstalk;ethylene;gibberellin;seed germination and dormancy
该方案是基于正文中引证的种子对乙烯、脱落酸或者GA响应的遗传分析、芯片数据和生理研究。乙烯通过抑制ABA的合成和促进它的失活或者分解代谢下调ABA的积累, 也负调控ABA信号。ABA通过ACS和ACO的活性抑制乙烯的生物合成。乙烯也增强GA的代谢和信号, 反过来也一样。“→”和“┤”分别表示信号级联的不同元素之间的正、负相互作用。根据Corbineau等[ 16]重绘。 Fig. 3Interaction among ethylene, abscisic acid, and gibberellin in the regulation of seed germination and dormancy
This scheme is based on genetic analyses, microarray data, and physiological studies on seed responsiveness to ethylene, ABA or GA cited in the text. Ethylene down-regulates ABA accumulation by both inhibiting its synthesis and promoting its inactivation or catabolism, and also negatively regulates ABA signaling. ABA inhibits ethylene biosynthesis through ACS and ACO activities. Ethylene also improves the GA metabolism and signaling, and vice versa. “→” and“ ┤” indicate positive and negative interactions between the different elements of the signaling cascade, respectively. Redrew from Corbineau et al. [ 16].
Xu HH, LiN, Liu SJ, Wang WQ, Wang WP, ZhangH, Cheng HY, Song SQ . Research progress in seed germination and its control Acta Agric Sin, 2014,40:1141-1156 (in Chinese with English abstract). [本文引用: 2]
Bewley JD, Bradford KJ, HilhorstH W M, NonogakiH . Physiology of Development, Germination and Dormancy, 3rd edn New York: Springer, 2013. pp 247-297. [本文引用: 1]
FinkelsteinR, ReevesW, AriizumiT, SreberC . Molecular aspects of seed dormancy Annu Rev Plant Biol, 2008,59:387-415. DOI: 10.1146/annurev.arplant.59.032607.092740 URL [本文引用: 1]
Song SQ. Seed dormancy. In: The Editorial Board of Agricultural Science for 10000 Selected Problems in Sciences, eds. 10000 Selected Problems in Sciences. Beijing: Science Press, 2011. pp 31-35(in Chinese). [本文引用: 1]
ShuK, Liu XD, XieQ, He ZH . Two faces of one seed: hormonal regulation of dormancy and germination Mol Plant, 2016,9:34-45. DOI: 10.1016/j.molp.2015.08.010 URL [本文引用: 2]
Hoang HH, SechetJ, BaillyC, LeymarieJ, CorbineauF . Inhibition of germination of dormant barley (Hordeum vulgare L.) grains by blue light as related to oxygen and hormonal regulation. Plant Cell Environ, 2014,37:1393-1403.
Lee HG, LeeK, Seo PJ . The Arabidopsis MYB96 transcription factors play a role in seed dormancy Plant Mol Biol, 2015,87:371-381. DOI: 10.1007/s11103-015-0283-4 URL [本文引用: 1]
ArcE, SechetJ, CorbineauF, RajjouL, Marion-PollA . ABA crosstalk with ethylene and nitric oxide in seed dormancy and germination Front Plant Sci, 2013,4:63. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.3389/fpls.2013.00063. [本文引用: 8]
LinkiesA, Leubner-MetzgerG . Beyond gibberellins and abscisic acid: how ethylene and jasmonates control seed germination Plant Cell Rep, 2012,31:253-270. DOI: 10.1007/s00299-011-1180-1 URL [本文引用: 4]
MiransariM, Smith DL . Plant hormones and seed germination Environ Exp Bot, 2014,99:110-121. DOI: 10.1016/j.envexpbot.2013.11.005 URL [本文引用: 3]
CorbineauF, XiaQ, BaillyC, EI-Maarouf-BouteauH . Ethylene, a key factor in the regulation of seed dormancy Front Plant Sci, 2014,5:539. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.3389/fpls.2014.00539. [本文引用: 17]
Khan NA, KhanM I R, FerranteA, PoorP . Editorial: Ethylene: a key regulatory molecule in plants Front Plant Sci, 2017,8:1782. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.3389/fpls.2017.01782. URL [本文引用: 1]
Khan NA, KhanM I R . The Ethylene: from senescence hormone to key player in plant metabolism J Plant Biochem Physiol, 2014,2:e124. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.4172/2329-9029.1000e124. [本文引用: 1]
LinZ, ZhongS, GriersonD . Recent advances in ethylene research J Exp Bot, 2009,60:3311-3336. DOI: 10.1093/jxb/erp204 URL [本文引用: 7]
Schaller GE . Ethylene and the regulation of plant development BMC Biol, 2012,10:9. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.1186/1741-7007-10-9. [本文引用: 1]
MüllerM, Munné-BoschS . Ethylene response factors: a key regulatory hub in hormone and stress signaling Plant Physiol, 2015,169:32-41. DOI: 10.1104/pp.15.00677 URL [本文引用: 2]
Thao NP, KhanM I R, ThuN B A, HoangX L T, AsgherM, KhanN A, TranL S P . Role of ethylene and its cross talk with other signaling molecules in plant responses to heavy metal stress Plant Physiol, 2015,169:73-84. DOI: 10.1104/pp.15.00663 URL [本文引用: 1]
Arraes F BM, Beneventi MA, de SaM E L, Paixao J FR, Albuquerque E VS, Marin S RR, PurgattoE, Nepomuceno AL, Grossi-de-Sa MF . Implications of ethylene biosynthesis and signaling in soybean drought stress tolerance BMC Plant Biol, 2015,15:213. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.1186/s12870-015-0597-z. URL [本文引用: 1]
Habben JE, BaoX, Bate NJ, DeBruinJ L, DolanD, HasegawaD, HelentjarisT G, LafitteR H, LovanN, MoH, ReimannK, SchusslerJ R . Transgenic alteration of ethylene biosynthesis increases grain yield in maize under field drought-stress conditions Plant Biotechnol J, 2014,12:685-693. DOI: 10.1111/pbi.2014.12.issue-6 URL [本文引用: 1]
LiebermanM, Mapson LW . Genesis and biogenesis of ethylene Nature, 1964,204:343-345. DOI: 10.1038/204343a0 [本文引用: 1]
LiebermanM, KunishiA, Mapson LW, Wardale DA . Stimulation of ethylene production in apple tissue slices by methionine Plant Physiol, 1966,41:76-82. [本文引用: 1]
Yang SF, Hoffman NE . Ethylene biosynthesis and its regulation in higher plants Annu Rev Plant Physiol, 1984,35:155-189. DOI: 10.1146/annurev.pp.35.060184.001103 URL [本文引用: 3]
Iglesias-FernandezR, MatillaA . Genes involved in ethylene and gibberellins metabolism are required for endosperm-limited germination of Sisymbrium officinale L. seeds. Planta, 2010,231:653-664. [本文引用: 3]
YamagamiT, TsuchisakaA, YamadaK, Haddon WF, Harden LA, TheologisA . Biochemical diversity among the 1-aminocyclopropane-1-carboxylate synthase isozymes encoded by the Arabidopsis gene family J Biol Chem, 2003,278:49102-49112. DOI: 10.1074/jbc.M308297200 URL [本文引用: 1]
TsuchisakaA, YuG, JinH, Alonso JM, Ecker JR, ZhangX, GaoS, TheologisA . A combinatorial interplay among the 1-aminocyclopropane-1-carboxylate isoforms regulates ethylene biosynthesis in Arabidopsis thaliana. Genetics, 2009,183:979-1003. [本文引用: 2]
Van de PoelB, Van Der StraetenD . 1-aminocyclopropane-1- carboxylic acid (ACC) in plants: more than just the precursor of ethylene! Front Plant Sci, 2014,5:640. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.3389/fpls.2014.%2000640. [本文引用: 3]
Yoon GM, KieberJ J . 1-Aminocyclopropane-1-carboxylic acid as a signalling molecule in plants AOB Plants, 2013, 5: plt017. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.1093/aobpla/plt017. [本文引用: 1]
Christians MJ, Gingerich DJ, HansenM, Binder BM, Kieber JJ, Vierstra RD . The BTB ubiquitin ligases ETO1, EOL1, and EOL2 act collectively to regulate ethylene biosynthesis in Arabidopsis by controlling type-2 ACC synthase levels Plant J, 2009,57:332-345. DOI: 10.1111/tpj.2009.57.issue-2 URL [本文引用: 1]
Lyzenga WJ, Booth JK, Stone SL . The Arabidopsis RING- type E3 ligase XBAT32 mediates the proteasomal degradation of the ethylene biosynthetic enzyme, 1-aminocyclopropane- 1-carboxylate synthase 7 Plant J, 2012,71:23-34. DOI: 10.1111/tpj.2012.71.issue-1 URL [本文引用: 1]
KamiyoshiharaY, IwataM, FukayaT, TatsukiM, MoriH . Turnover of LeACS2, a wound-inducible 1-aminocyclopropane- 1-carboxylic acid synthase in tomato, is regulated by phosphorylation/dephosphorylation Plant J, 2010,64:140-150. [本文引用: 1]
LudwikowA, CieslaA, Kasprowicz-MaluskiA, MitulaF, TajdelM, GalganskiL, Ziolkowski PA, KubiakP, MaleckA, PiechalakA, SzabatM, GorskaA, DabrowskiM, IbragimowI, SadoqskiJ . Arabidopsis protein phosphatase 2C ABI1 interacts with type I ACC synthases and is involved in the regulation of ozone-induced ethylene biosynthesis Mol Plant, 2014,7:960-967. DOI: 10.1093/mp/ssu025 URL [本文引用: 1]
Skottke KR, Yoon GM, Kieber JJ, DeLongA . Protein phosphatase 2A controls ethylene biosynthesis by differentially regulating the turnover of ACC synthase isoforms PLoS Genet, 2011,7:e1001370. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.1371/journal.pgen.10%2001370. URL [本文引用: 1]
Van de PoelB, BulensI, MarkoulaA, HertogM L A T M, DreesenR, WirtzM, VandoninckS, OppermannY, KeulemansJ, HellR, WaelkensE, De Proft MP, SauterM, Nicolai BM, Geeraerd AH . Targeted systems biology profiling of tomato fruit reveals coordination of the Yang cycle and a distinct regulation of ethylene biosynthesis during postclimacteric ripening Plant Physiol, 2012,160:1498-1514. DOI: 10.1104/pp.112.206086 URL [本文引用: 1]
Matilla AJ, Matilla-VazquezM A . Involvement of ethylene in seed physiology Plant Sci, 2008,175:87-97. DOI: 10.1016/j.plantsci.2008.01.014 URL [本文引用: 3]
VerveridisP, JohnP . Complete recovery in vitro of ethylene- forming enzyme-activity Phytochemistry, 1991,30:725-727. DOI: 10.1016/0031-9422(91)85241-Q URL [本文引用: 1]
Murphy LJ, Robertson KN, Harroun SG, Brosseau CL, Werner-ZwanzigerU, MoilanenJ, Tuononen HM, ClyburneJ A C . A simple complex on the verge of breakdown: isolation of the elusive cyanoformate ion Science, 2014,344:75-78. DOI: 10.1126/science.1250808 URL [本文引用: 2]
叶永健, 宋松泉 . Fe 2+和CO2对番木瓜ACC氧化酶活性的影响 中山大学学报, 1997,36(2):18-21. [本文引用: 1]
Ye YJ, Song SQ . Effect of Fe 2+ and CO2 on 1-aminocyclopropane-1-carboxylate oxidase from papaya (Carica papaya L.) fruit. Acta Sci Nat Univ Sunyatseni, 1997,36(2):18-21. [本文引用: 1]
Dong JG, Fernandezmaculet JC, Yang SF . Purification and characterization of 1-aminocyclopropane-1-carboxylate oxidase from apple fruit Proc Natl Acad Sci USA, 1992,89:9789-9793. DOI: 10.1073/pnas.89.20.9789 URL [本文引用: 1]
Song SQ, Ye YJ . Effect of O2, ACC and CO2 concentration on the activity of partially purified ACC oxidase from papaya Acta Phytophysiol Sin, 2001,27:387-392. [本文引用: 1]
Hudgins JW, Ralph SG, Franceschi VR, BohlmannJ . Ethylene in induced conifer defense: cDNA cloning, protein expression, and cellular and subcellular localization of 1-aminocyclopropane- 1-carboxylate oxidase in resin duct and phenolic parenchyma cells Planta, 2006,224:865-877. DOI: 10.1007/s00425-006-0274-4 URL [本文引用: 1]
RamassamyS, OlmosE, BouzayenM, Pech JC, LatcheA . 1-aminocyclopropane-1-carboxylate oxidase of apple fruitis periplasmic J Exp Bot, 1998,49:1909-1915. [本文引用: 1]
De PaepeA, VuylstekeM, vanHummelen P, ZabeauM, van Der StaetenD . Transcriptional profiling by cDNA-AFLP and microarray analysis reveals novel insights into the early response to ethylene in Arabidopsis Plant J, 2004,39:537-559. DOI: 10.1111/tpj.2004.39.issue-4 URL [本文引用: 1]
Van de PoelB, BulensI, HertogM L A T M, NicolaiB, GeeraerdA . A transcriptomics-based kinetic model for ethylene biosynthesis in tomato (Solanum lycopersicum) fruit: development, validation and exploration of novel regulatory mechanisms. New Phytol, 2014,202:952-963. [本文引用: 1]
Binder BM, ChangC, Schaller GE . Perception of ethylene by plants-ethylene receptors Annu Plant Rev, 2012,44:117-145. [本文引用: 2]
Stepanova AN, Alonso JM . Ethylene signaling and response: where different regulatory modules meet Curr Opin Plant Biol, 2009,12:548-555. DOI: 10.1016/j.pbi.2009.07.009 URL [本文引用: 4]
BakshiA, PiyaS, Fernandez JC, ChervinC, HeweziT, Bindera BM . Ethylene receptors signal via a noncanonical pathway to regulate abscisic acid responses Plant Physiol, 2018,176:910-929. DOI: 10.1104/pp.17.01321 URL [本文引用: 1]
JuC, ChangC . Advances in ethylene signalling: protein complexes at the endoplasmic reticulum membrane. AOB Plants, 2012, 2012: pls031. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.1093/aobpla/pls031. [本文引用: 2]
Shakeel SN, WangX, Binder BM, SchallerG E . Mechanisms of signal transduction by ethylene: overlapping and non-overlapping signalling roles in a receptor family AOB Plants, 2013, 5: plt010. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.1093/aobpla/plt010. [本文引用: 1]
Chen YF, GaoZ, Kerris RJ, WangW, Binder BM, Schaller GE . Ethylene receptors function as components of high-molecular- mass protein complexes in Arabidopsis PLoS One, 2010,5:e8640. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.1371/journal.pone.0008640. URL [本文引用: 1]
HirayamaT, Kieber JJ, HirayamaN, KoganM, GuzmanP, NourizadehS, Alonso JM, Dailey WP, DancisA, Ecker JR . RESPONSIVE-TO-ANTAGONIST1, a Menkes/Wilson disease related copper transporter, is required for ethylene signaling in Arabidopsis Cell, 1999,97:383-393. DOI: 10.1016/S0092-8674(00)80747-3 URL [本文引用: 2]
Binder BM, Rodriguez FI, Bleecker AB . The copper transporter RAN1 is essential for biogenesis of ethylene receptors in Arabidopsis J Biol Chem, 2010,285:37263-37270. DOI: 10.1074/jbc.M110.170027 URL [本文引用: 1]
Resnick JS, Wen CK, Shockey JA, ChangC . REVERSION-TO ETHYLENE SENSITIVITY 1, a conserved gene that regulates ethylene receptor function in Arabidopsis Proc Natl Acad Sci USA, 2006,103:7917-7922. DOI: 10.1073/pnas.0602239103 URL [本文引用: 1]
Dong CH, RivarolaM, Resnick JS, Maggin BD, ChangC . Subcellular colocalization of Arabidopsis RTE1 and ETR1 supports a regulatory role for RTE1 in ETR1 ethylene signaling Plant J, 2008,53:275-286. [本文引用: 1]
Resnick JS, RivarolaM, ChangC . Involvement of RTE1 in conformational changes promoting ETR1 ethylene receptor signaling in Arabidopsis Plant J, 2008,56:423-431. DOI: 10.1111/tpj.2008.56.issue-3 URL [本文引用: 1]
MayerhoferH, PanneerselvamS, Mueller-DieckmannJ . Protein kinase domain of CTR1 from Arabidopsis thaliana promotes ethylene receptor cross talk. J Mol Biol, 2012,415:768-779. [本文引用: 1]
Alonso JM, HirayamaT, RomanG, NourizadehS, Ecker JR . EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis Science, 1999,284:2148-2152. DOI: 10.1126/science.284.5423.2148 URL [本文引用: 1]
WenX, ZhangC, JiY, ZhaoQ, HeW, AnF, JiangL, GuoH . Activation of ethylene signaling is mediated by nuclear translocation of the cleaved EIN2 carboxyl terminus Cell Res, 2012,22:1613-1616. DOI: 10.1038/cr.2012.145 [本文引用: 3]
Cho YH, LeeS, Yoo SD . EIN2 and EIN3 in ethylene signaling Annu Plant Rev, 2012,44:169-187. [本文引用: 1]
Bisson M MA, GrothG . New insight in ethylene signaling: autokinase activity of ETR1 modulates the interaction of receptors and EIN2 Mol Plant, 2010,3:882-889. DOI: 10.1093/mp/ssq036 URL [本文引用: 1]
AnF, ZhaoQ, JiY, LiW, JiangZ, YuX, ZhangC, HanY, HeW, LiuY, ZhangS, Ecker JR, GuoH . Ethylene-induced stabilization of ETHYLENE INSENSITIVE 3 and EIN3-LIKE1 is mediated by proteasomal degradation of EIN3 binding F-box 1 and 2 that requires EIN2 in Arabidopsis Plant Cell, 2010,22:2384-2401. DOI: 10.1105/tpc.110.076588 URL [本文引用: 2]
JuC, Yoon GM, Shemansky JM, Lin DY, Ying ZI, ChangJ, Garrett WM, KessenbrockM, GrothG, Tucker ML, CooperB, Kieber JJ, ChangC . CTR1 phosphorylates the central regulator EIN2 to control ethylene hormone signaling from the ER membrane to the nucleus in Arabidopsis Proc Natl Acad Sci USA, 2012,109:19486-19491. DOI: 10.1073/pnas.1214848109 URL [本文引用: 4]
QiaoH, ShenZ, Huang SC, Schmitz RJ, Urich MA, Briggs SP, Ecker JR . Processing and subcellular trafficking of ER-tethered EIN2 control response to ethylene gas Science, 2012,338:390-393. DOI: 10.1126/science.1225974 URL [本文引用: 2]
JiY, GuoH . From endoplasmic reticulum (ER) to nucleus: EIN2 bridges the gap in ethylene signaling Mol Plant, 2013,6:11-14. DOI: 10.1093/mp/sss150 URL [本文引用: 1]
LiJ, LiZ, TangL, YangY, ZouineM, BouzayenM . A conserved phosphorylation site regulates the transcriptional function of ETHYLENE-INSENSITIVE3-like1 in tomato J Exp Bot, 2012,63:427-439. DOI: 10.1093/jxb/err289 URL [本文引用: 1]
Chang KN, ZhongS, Weirauch MT, HonG, PelizzolaM, LiH, HuangS S C, SchmitzR J, UrichM A, KuoD, NeryJ R, QiaoH, YangA, JamaliA, ChenH, IdekerT, RenB, Bar-JosephZ, HughesT R, EckerJ R . Temporal transcriptional response to ethylene gas drives growth hormone cross-regulation in Arabidopsis eLife, 2013,2:e00675. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.7554/eLife.00675. URL [本文引用: 1]
Chen YF, Shakeel SN, BowersJ, Zhao XC, EtheridgeN, Schaller GE . Ligand-induced degradation of the ethylene receptor ETR2 through a proteasome-dependent pathway in Arabidopsis J Biol Chem, 2007,282:24752-24758. DOI: 10.1074/jbc.M704419200 URL [本文引用: 1]
Gagne JM, SmalleJ, Gingerich DJ, Walker JM, Yoo SD, YanagisawaS, Vierstra RD . Arabidopsis EIN3-binding F-box 1 and 2 form ubiquitin-protein ligases that repress ethylene action and promote growth by directing EIN3 degradation Proc Natl Acad Sci USA, 2004,101:6803-6808. DOI: 10.1073/pnas.0401698101 URL [本文引用: 2]
QiaoH, Chang KN, YazakiJ, Ecker JR . Interplay between ethylene, ETP1/ETP2 F-box proteins, and degradation of EIN2 triggers ethylene responses in Arabidopsis Genes Dev, 2009,23:512-521. DOI: 10.1101/gad.1765709 URL [本文引用: 3]
PotuschakT, LechnerE, ParmentierY, YanagisawaS, GravaS, KonczC, GenschikP . EIN3-dependent regulation of plant ethylene hormone signaling by two Arabidopsis F box proteins: EBF1 and EBF2 Cell, 2003,115:679-689. DOI: 10.1016/S0092-8674(03)00968-1 URL [本文引用: 1]
KonishiM, YanagisawaS . Ethylene signaling in Arabidopsis involves feedback regulation via the elaborate control of EBF2 expression by EIN3 Plant J, 2008,55:821-831. DOI: 10.1111/tpj.2008.55.issue-5 URL [本文引用: 1]
Kevany BM, Tieman DM, Taylor MG, Cin VD, Klee HJ . Ethylene receptor degradation controls the timing of ripening in tomato fruit Plant J, 2007,51:458-467. DOI: 10.1111/j.1365-313X.2007.03170.x URL [本文引用: 1]
Fu JR, Yang SF . Release of heat pretreatment-induced dormancy in lettuce seeds by ethylene or cytokinin in relation to the production of ethylene and the synthesis of 1-aminocyclopropane-1- carboxylic acid during germination J Plant Growth Regul, 1983,2:185-192. DOI: 10.1007/BF02042247 URL [本文引用: 1]
SiriwitayawanG, Geneve RL, Downie AB . Seed germination of ethylene perception mutants of tomato and Arabidopsis Seed Sci Res, 2003,13:303-314. DOI: 10.1079/SSR2003147 URL [本文引用: 1]
K?pczyńskiJ, SznigirP . Participation of GA3, ethylene, NO and HCN in germination of Amaranthus retroflexus L. seeds with various dormancy levels. Acta Physiol Plant, 2014,36:1463-1472. [本文引用: 3]
Van dePoel, SmetD, Van Der StraetenD . Ethylene and hormonal cross talk in vegetative growth and development Plant Physiol, 2015,169:61-72. DOI: 10.1104/pp.15.00724 URL [本文引用: 1]
Wood LA, Kester ST, Geneve RL . The physiological basis for ethylene-induced dormancy release in three Echinacea species with special reference to the influence of the integumentary tapetum. Sci Hortic, 2013: 156:63-72. [本文引用: 1]
LinY, YangL, PaulM, ZuY, TangZ . Ethylene promotes germination of Arabidopsis seed under salinity by decreasing reactive oxygen species: evidence for the involvement of nitric oxide simulated by sodium niroprusside Plant Physiol Biochem, 2013,73:211-218. DOI: 10.1016/j.plaphy.2013.10.003 URL [本文引用: 1]
Silva PO, Medina EF, Barros RS, Ribeiro DM . Germination of salt-stressed seeds as related to ethylene biosynthesis ability in three Stylosanthes species. J Plant Physiol, 2014,171:14-22. [本文引用: 1]
SinskaI . Interaction of ethephon with cytokinin and gibberellin during the removal of apple seed dormancy and germination of embryos Plant Sci, 1989,64:39-44. DOI: 10.1016/0168-9452(89)90149-0 URL [本文引用: 1]
CorbineauF, C?meD. Germination of sunflower seeds as related to ethylene synthesis and sensitivity: an overview. In: Vendrell M, Klee H, Pech J C, Romojaro F, eds. Biology and Biotechnology of the Plant Hormone Ethylene III. Amsterdam: IOS Press, 2003. pp 216-221. [本文引用: 3]
Ribeiro DM, Barros RS . Sensitivity to ethylene as a major component in the germination of seeds of Stylosanthes humilis. Seed Sci Res, 2006,16:37-45. [本文引用: 1]
GniazdowskaA, KrasuskaU, BogatekR . Dormancy removal in apple embryos by nitric oxide or cyanide involves modifications in ethylene biosynthetic pathway Planta, 2010,232:1397-1407. DOI: 10.1007/s00425-010-1262-2 URL [本文引用: 1]
ArgyrisJ, DahalP, HayashiE, Still DW, Bradford KJ . Genetic variation for lettuce seed thermoinhibition is associated with temperature-sensitive expression of abscisic acid, gibberellin, and ethylene biosynthesis, metabolism, and response genes Plant Physiol, 2008,148:926-947. DOI: 10.1104/pp.108.125807 URL [本文引用: 2]
HermannK, MeinhardJ, DobrevP, LinkiesA, PesekB, HessB, Machá?ková IM, FischerU, Leubner-MetzgerG . 1-Amynocyclopropane-1-carboxylic acid and abscisic acid during the germination of sugar beet (Beta vulgaris L.): a comparative study of fruits and seeds. J Exp Bot, 2007,58:3047-3060. [本文引用: 1]
KrasuskaU, CiackaK, DebskaK, BogatekR, GniazdowskaA . Dormancy alleviation by NO or HCN leading to decline of protein carbonylation levels in apple (Malus domestica Borkh.) embryos. J Plant Physiol, 2014,171:1132-1141. [本文引用: 1]
OraczK, El-Maarouf-BouteauH, BogatekR, CorbineauF, BaillyC . Release of sunflower seed dormancy by cyanide: cross-talk with ethylene signaling pathway J Exp Bot, 2008,59:2241-2251. DOI: 10.1093/jxb/ern089 URL [本文引用: 2]
Chiwocha S DS, Cutler AJ, Abrams SR, Ambrose SJ, YangJ, Kermode AR . The ert1-2 mutation in Arabidopsis thaliana affects the abscisic acid, auxin, cytokinin and gibberellin metabolic pathways during main tenance of seed dormancy, moist-chilling and germination. Plant J, 2005,42:35-48. [本文引用: 5]
SubbiahV, Reddy KJ . Interactions between ethylene, abscisisc acid and cytokinin during germination and seedling establishment in Arabidopsis J Biosci, 2010,35:451-458. DOI: 10.1007/s12038-010-0050-2 URL [本文引用: 3]
Jimenez JA, RodriguezD, Calvo AP, Mortensen LC, NicolasG, NicolasC . Expression of a transcription factor (FsERF1) involved in ethylene signaling during the breaking of dormancy in Fagus sylvatica seeds. Physiol Plant, 2005,125:373-380. [本文引用: 1]
Cadman C SC, Toorop PE, Hilhorst H WM, Finch-Savage WE . Gene expression profiles of Arabidopsis Cvi seeds during dormancy cycling indicate a common underlying dormancy control mechanism Plant J, 2006,46:805-822. DOI: 10.1111/tpj.2006.46.issue-5 URL [本文引用: 1]
LinkiesA, MüllerK, MorrisK, Ture?kováV, WenkM, CadmanC S C, CorbineauF, StrnadM, LynnJ R, Finch-SavageW E, Leubner-MetzgerG . Ethylene interacts with abscisic acid to regulate endosperm rupture during germination: a comparative approach using Lepidium sativum and Arabidopsis thaliana. Plant Cell, 2009,21:3803-3822. [本文引用: 6]
NonogakiH . Seed dormancy and germination-emerging mechanism and new hypotheses Front Plant Sci, 2014,e5:233. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.3389/fpls.2014.00233. [本文引用: 2]
LiuX, Hou XL . Antagonistic regulation of ABA and GA in metabolism and signaling pathways Front Plant Sci, 2018,9:251. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.3389/fpls.2018.00251. URL [本文引用: 1]
Dong TT, Tong JH, Xiao LT, Cheng HY, Song SQ . Nitrate, abscisic acid and gibberellin interactions on the thermoinhibition of lettuce seed germination Plant Growth Regul, 2012,66:191-202. DOI: 10.1007/s10725-011-9643-5 URL [本文引用: 1]
DongZ, YuY, LiS, WangJ, TangS, HuangR . Abscisic acid antagonizes ethylene production through the ABI4-mediated transcriptional repression of ACS4 and ACS8 in Arabidopsis. Mol Plant, 2016,9:126-135. [本文引用: 2]
PenfieldS, LiY, Gilday AD, GrahamS, Graham IA . Arabidopsis ABA INSENSITIVE 4 regulates lipid mobilization in the embryo and reveals repression of seed germination by the endosperm Plant Cell, 2006,18:1887-1899. DOI: 10.1105/tpc.106.041277 URL [本文引用: 2]
Cheng WH, Chiang MH, Hwang SG, Lin PC . Antagonism between abscisic acid and ethylene in Arabidopsis acts inparallel with the reciprocal regulation of their metabolism and signaling pathways Plant Mol Biol, 2009,71:61-80. DOI: 10.1007/s11103-009-9509-7 URL [本文引用: 3]
Alonso-RamirezA, RodriguezD, ReyesD, Jimenez JA, NicolasG, NicolasC . Functional analysis in Arabidopsis of FsPTP1, a tyrosine phosphatase from beechnuts, reveals its role as a negative regulator of ABA signaling and seed dormancy and suggests its involvement in ethylene signaling modulation. Planta, 2011,234:589-597. [本文引用: 1]
Calvo AP, NicolasC, LorenzoO, NicolasG, RodriguezD . Evidence for positive regulation by gibberellins and ethylene of ACC oxidase expression and activity during transition from dormancy to germination in Fagus sylvatica L. seeds. J Plant Growth Regul, 2004,23:44-53. [本文引用: 2]
Calvo AP, NicolasC, NicolasG, RodriguezD . Evidence of a crosstalk regulation of a GA 20-oxidase (FsGA20ox1) by gibberellins and ethylene during the breaking of dormancy in Fagus sylvatica seeds. Physiol Plant, 2004,120:623-630. [本文引用: 2]
LorenzoO, RodriguezD, NicolasC, NicolasG. Characterization and expression of two protein kinase and an EIN3-like genes, which are regulated by ABA and GA3 in dormant Fagus sylvatica seeds. In: Black M, Bradford K J, Vazquez-Ramos J, eds. Seed Biology: Advances and Applications. Wallingford: CAB International, 2000. pp 329-340. [本文引用: 1]
DillA, Thomas SG, Steber CM, Sun TP . The Arabidopsis F-box protein SLEEPY1 targets gibberellin signaling repressors for gibberellin-induced degradation Plant Cell, 2004,16:1392-1405. DOI: 10.1105/tpc.020958 URL [本文引用: 1]
AchardP, BaghourM, ChappleA, HeddenP, VanDer Straeten D, GenschikP, MoritzT, HarberdN P . The plant stress hormone ethylene controls floral transition via DELLA-dependent regulation of floral meristem-identity genes Proc Natl Acad Sci USA, 2007,104:6484-6489. DOI: 10.1073/pnas.0610717104 URL [本文引用: 1]
SchwechheimerC . Understanding gibberellic acid signaling: are we there yet? Curr Opin Plant Biol, 2008,11:9-15. DOI: 10.1016/j.pbi.2007.10.011 URL [本文引用: 1]
ZhangJ, YuJ, WenC-K . An alternate route of ethylene receptor signaling Front Plant Sci, 2014,5:648. doi: http://zwxb.chinacrops.org/article/2019/0496-3490/10.3389/fpls.2014.00648. [本文引用: 1]
NambaraE, OkamotoM, TatematsuK, YanoR, SeoM, KamiyaY . Abscisic acid and the control of seed dormancy and germination Seed Sci Res, 2010,20:55-67. DOI: 10.1017/S0960258510000012 URL [本文引用: 1]
Liu SJ, Song SH, Wang WQ, Song SQ . De novo assembly and characterization of germinating lettuce seed transcriptome using Illumina paired-end sequencing Plant Physiol Biochem, 2015,96:154-162. DOI: 10.1016/j.plaphy.2015.07.020 URL [本文引用: 1]
Wang WQ, Song BY, Deng ZJ, WangY, Liu SJ, M?ller IM, Song SQ . Proteomic analysis of Lactuca sativa seed germination and thermoinhibition by sampling of individual seeds at germination and removal of storage proteins by PEG fractionation. Plant Physiol, 2015,167:1332-1350. [本文引用: 1]
Xu HH, Liu SJ, Song SH, Wang WQ, M?ller IM, Song SQ . Proteome changes associated with dormancy release of Dongxiang wild rice seeds J Plant Physiol, 2016,206:68-86. DOI: 10.1016/j.jplph.2016.08.016 URL [本文引用: 1]
 ,1,3,*, 刘军2, 徐恒恒2, 张琪2, 黄荟3, 伍贤进3 1 中国科学院植物研究所, 北京100093
,1,3,*, 刘军2, 徐恒恒2, 张琪2, 黄荟3, 伍贤进3 1 中国科学院植物研究所, 北京100093
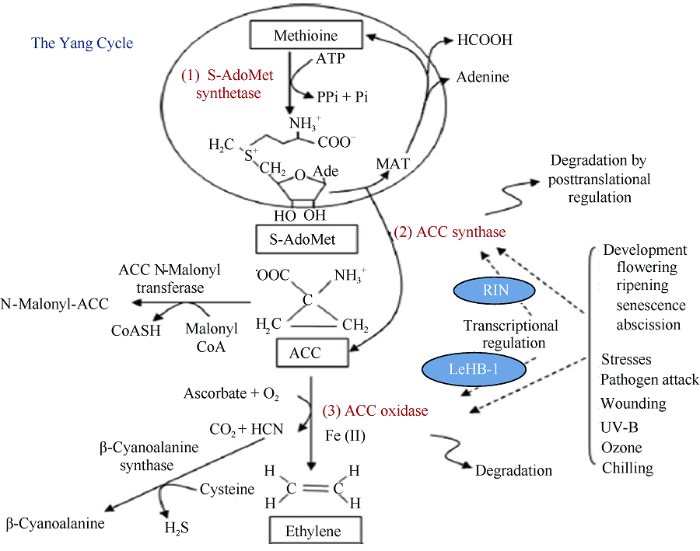 新窗口打开| 下载原图ZIP| 生成PPT
新窗口打开| 下载原图ZIP| 生成PPT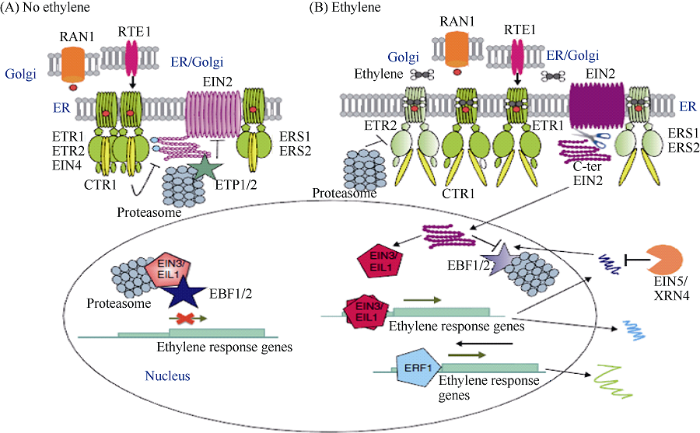 新窗口打开| 下载原图ZIP| 生成PPT
新窗口打开| 下载原图ZIP| 生成PPT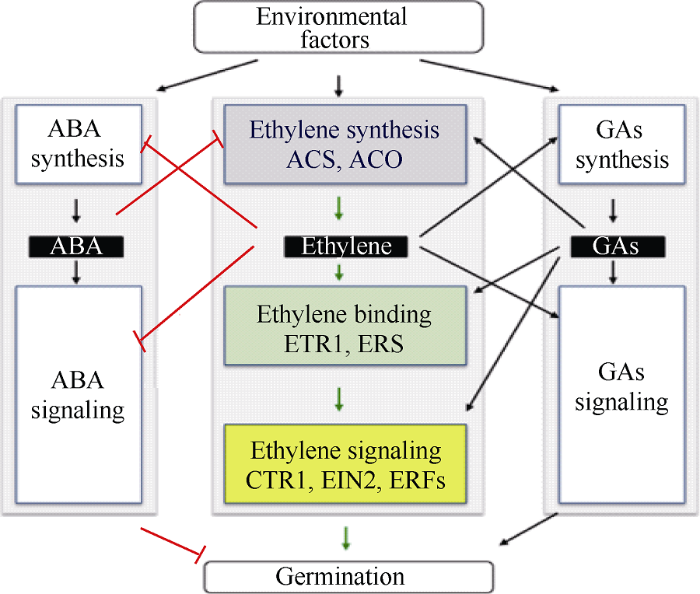 新窗口打开| 下载原图ZIP| 生成PPT
新窗口打开| 下载原图ZIP| 生成PPT