与延伸起始因子4E结合的肽和化合物的制作方法
与延伸起始因子4e结合的肽和化合物
1.对相关申请的交叉引用
2.本技术要求于2019年3月29日提交的第10201902892r号新加坡申请的优先权,其内容通过引用整体并入本文以用于所有目的。
技术领域
3.本发明涉及细胞增殖抑制肽。特别是,本发明涉及与延伸因子4e(eif4e)结合的肽。
背景技术:
4.最常发生的癌症突变是在进入翻译机制的信号转导通路中发现的。这些包括经过充分研究和验证的癌基因,例如myc、ras和pik3c。此外,通常发现广泛的翻译起始因子在肿瘤中被扩增或被错误调节。eif4e(延伸起始因子4e)在大多数癌症中经常被错误调节和超表达,并且与一大类致癌相关蛋白(例如vegf和c
‑
myc)的上调密切相关。
5.eif4e通过直接与mrna的5'帽结构相互作用,在翻译起始中发挥重要作用。所有核转录的mrna都具有这种5'帽结构(m7gtp),该结构由在第7位甲基化并通过5'到5'三磷酸桥连接到mrna的第一个核苷酸的鸟苷组成。包括癌基因在内的绝大多数真核mrna都以eif4e帽依赖性方式翻译,eif4e的超表达导致这些基因的翻译增加。因此,需要控制eif4e活性平以防止不受控制的增殖。
6.调节eif4e水平和活性的一种方式是通过eif4e结合蛋白(4e
‑
bp)调节eif4e活性。4e
‑
bp负向调节eif4e,并与eif4g共享共同的结合基序,它们利用该基序直接结合eif4e并竞争性置换eif4g支架蛋白。eif4g的置换损害了eif4f复合物在5'帽结构上的组装,并阻碍了帽依赖性翻译。
7.因此,需要鉴定具有不同于上述作用机制的eif4e结合剂,以规避新兴抗性机制,已针对靶向参与调节蛋白质合成的eif4e上游信号通路的化合物报道该新兴抗性机制。还需要开发用于抗癌治疗的eif4e结合剂。
8.发明概述
9.一方面,提供了包含氨基酸序列cex1gx2x3x4x5c(seq id no:1)的肽,其中x1是选自由苏氨酸(t)、甲硫氨酸(m)或亮氨酸(l)组成的组的氨基酸;其中x2是选自由疏水性氨基酸、芳香族氨基酸、经修饰的疏水性氨基酸和经修饰的芳香族氨基酸组成的组的氨基酸;其中x3是选自由疏水性氨基酸、芳香族氨基酸、经修饰的疏水性氨基酸和经修饰的芳香族氨基酸组成的组的氨基酸;其中x4是任何氨基酸;其中x5是任何氨基酸;其中两个半胱氨酸残基通过二硫键连接;并且其中肽与延伸起始因子4e(eif4e)结合。
10.在另一方面,提供了本文所述的肽作为药物的用途。
11.在另一方面,提供本文所述的肽在治疗与帽依赖性翻译失调相关的疾病状况中的用途。
12.在另一方面,提供了包含本文所述的肽和药学上可接受的载体的药物组合物。
13.在另一方面,提供了本文所述的肽或本文所述的药物组合物在制造用于治疗与帽依赖性翻译失调相关的疾病状况的药物中的用途。
14.在另一方面,提供了治疗有需要的个体的与帽依赖性翻译失调相关的疾病状况的方法,包括向个体施用本文所述的肽或本文所述的药物组合物。
15.定义
16.如本文所用,术语“蛋白质”或“肽”是指氨基酸的聚合形式。蛋白质应理解为比肽包含更多的氨基酸,并且“蛋白质”通常包含至少约60个氨基酸,而肽通常包含2个至约60个氨基酸。蛋白质或肽中的氨基酸可以以d或l构型或两种构型的组合存在,并且可以进行化学或生物化学修饰。氨基酸的修饰可发生在侧链或连接氨基酸的肽键处。可以修饰肽键(
‑
conh
‑
)以改变肽键的特性。也可以通过添加共价结合的化学基团,例如生物素、硫醇、半胱氨酸、酰胺、羧基、直链或支链烷基、伯胺或仲胺、脂质、磷脂、脂肪酸、胆固醇等,对肽进行化学修饰。这些化学基团可以添加在肽的n
‑
末端和/或c
‑
末端或肽内。也可以通过在n
‑
和c
‑
末端之间在肽内添加一个或多个其他键,例如二硫键,来修饰肽。也可以化学修饰n
‑
末端的胺基,例如通过乙酰化添加帽。
17.如本文所用,术语“氨基酸”是指包含胺(
‑
nh2)和羧基(
‑
cooh)官能团和侧链的分子。氨基酸可由核酸的三联密码子(规范氨基酸)或变异密码子(非规范氨基酸)编码。氨基酸可以表征为α
‑
氨基酸或β
‑
氨基酸。在α
‑
氨基酸中,胺基和羧基连接于第一个碳。在β
‑
氨基酸中,胺基和羧基相连于相邻的碳。氨基酸还可以表征为d
‑
氨基酸(其中氨基的立体α碳具有d
‑
构型)或l
‑
氨基酸(其中氨基的立体α碳具有l
‑
构型)。氨基酸也可根据其侧链的性质进行分类。例如,氨基酸可分为酸性、碱性、极性、疏水性或芳香性,并且氨基酸可归入一个或多个类别。芳香族氨基酸包括苯丙氨酸、色氨酸、酪氨酸和组氨酸。疏水性氨基酸包括甘氨酸(gly)、丙氨酸(ala)、缬氨酸(val)、亮氨酸(leu)、异亮氨酸(ile)、脯氨酸(pro)、苯丙氨酸(phe)、甲硫氨酸(met)和色氨酸(trp)。极性氨基酸包括丝氨酸(ser)、苏氨酸(thr)、半胱氨酸(cys)、天冬酰胺(asn)、谷氨酰胺(gln)和酪氨酸(tyr)。碱性氨基酸包括精氨酸(arg)、赖氨酸(lys)和组氨酸(his),酸性氨基酸包括天冬氨酸(aspartic acid)或天冬氨酸(aspartate)(asp)和谷氨酸(glutamic acid)或谷氨酸(glutamate)(glu)。
18.本文所用的氨基酸可以是天然存在的、非天然存在的或合成的。也可以通过取代侧链上一个或多个基团来修饰本文所用的氨基酸。
19.表1:氨基酸
20.[0021][0022]
本文在经修饰的氨基酸的语境中所用的术语“经修饰的”、“修饰”或其语法变体是指侧链上的一个或多个基团被取代的氨基酸。可以用以下基团取代一个或多个基团:烷基、烯基、炔基、硫代烷基、环烷基、环烯基、杂环烷基、卤素、羧基、卤代烷基、卤代炔基、羟基、烷氧基、硫代烷氧基、烯氧基、卤代烷氧基、卤代烯氧基、硝基、氨基、硝基烷基、硝基烯基、硝基炔基、硝基杂环基、烷基氨基、二烷基氨基、烯基氨基、炔基氨基、酰基、烯酰基、炔酰基、酰氨基、二酰基氨基、酰氧基、烷基磺酰氧基、杂环氧基、杂环氨基、卤代杂环烷基、烷基亚磺酰基、烷基羰基氧、烷硫基、酰硫基、含磷基团例如膦羧基和氧膦基、芳基、杂芳基、烷基芳基、烷基杂芳基、氰基、氰酸酯、异氰酸酯、
‑
c(o)nh(烷基)和
‑
c(o)n(烷基)2。
[0023]
本文所用术语“烷基”在其含义内包括具有1
‑
10个碳原子,例如具有1、2、3、4、5、6、7、8、9或10个碳原子的一价(“烷基”)和二价(“亚烷基”)直链或支链饱和脂肪族基团。
[0024]
术语“烯基”在其含义内包括具有2
‑
10个碳原子,例如具有2、3、4、5、6、7、8、9或10个碳原子,并且如适用在烷基链中的任何位置具有至少一个e、z、顺式或反式立体化学双键的一价(“烯基”)和二价(“亚烯基”)直链或支链不饱和脂肪族烃基团。
[0025]
本文所用术语“炔基”在其含义内包括具有2
‑
10个碳原子并且在碳链任何位置具有至少一个三键的一价(“炔基”)和二价(“亚炔基”)直链或支链不饱和脂肪族烃基团。
[0026]
本文所用术语“环烷基”是指环状饱和脂肪族基团,并且在其含义内包括具有3
‑
10碳原子,例如具有3、4、5、6、7、8、9或10个碳原子的一价(“环烷基”)和二价(“环亚烷基”)饱和、单环、双环、多环或稠合多环烃基。
[0027]
本文所用术语“环烯基”是指环状不饱和脂肪族基团,并且在其含义内包括具有3
‑
10个碳原子且如适用在烷基链的任何位置具有至少一个e、z、顺式或反式立体化学双键的一价(环烯基)和二价(环亚烯基)的单环、双环、多环或稠合多环烃基。环烯基的实例包括但不限于环丙烯基、环戊烯基、环己烯基等。
[0028]
本文所用术语“杂环烷基”在其含义内包括具有3
‑
10个环原子的一价(“杂环烷基”)和二价(“杂环亚烷基”)饱和、单环、双环、多环或稠合烃基,其中1
‑
5个环原子是选自o、n、nh或s的杂原子。实例包括吡咯烷基、哌啶基、奎宁环基、氮杂环丁烷基、吗啉基、四氢噻吩基、四氢呋喃基、四氢吡喃基等。
[0029]
本文所用术语“杂环烯基”在其含义内包括具有3
‑
10个环原子并具有至少1个双键的一价(杂环烯基)和二价(杂环亚烯基)饱和、单环、双环、多环或稠合烃基,其中1
‑
5个环原子是选自o、n、nh或s的杂原子。
[0030]
本文所用术语“杂芳香族基团(heteroaromatic group)”和诸如“杂芳基(heteroaryl)”或“杂亚芳基(heteroarylene)”等变体在其含义内包括具有6
‑
20个原子的一价(“杂芳基”)和二价(“杂亚芳基”)单核、多核、缀合和稠合芳基,其中1
‑
6个原子是选自o、n、nh和s的杂原子。
[0031]
本文所用术语“杂原子”或变体例如“杂
‑
(hetero
‑
)”是指o、n、nh和s。
[0032]
本文所用术语“烷氧基”是指直链或支链烷氧基。
[0033]
本文在取代的语境中所用的术语“氨基”是指
–
nr
a
r
b
形式的基团,其中r
a
和r
b
分别选自包括但不限于氢、任选取代的烷基、任选取代的烯基、任选取代的炔基和任选取代的芳基。
[0034]
本文所用术语“芳香族基团(aromatic group)”或诸如“芳基(aryl)”或“亚芳基(arylene)”等变体是指具有6
‑
10个碳原子的一价(芳基)和二价(亚芳基)单核、多核、缀合和稠合残基的芳香族烃。
[0035]
本文所用术语“芳烷基”在其含义内包括连接于二价饱和、直链和支链亚烷基的一价(“芳基”)和二价(“亚芳基”)单核、多核、缀合和稠合芳香族烃基。
[0036]
本文所用术语“杂芳烷基”在其含义内包括连接于二价饱和、直链和支链亚烷基的一价(“杂芳基”)和二价(“杂亚芳基”)单核、多核、缀合和稠合芳香族烃基。
[0037]
本文所用术语“卤素(halogen)”或诸如“卤化物”或“卤代(halo)”等变体是指氟、氯、溴、碘和氰卤化物。
[0038]
本文所用术语“平衡解离常数”或“解离常数”(k
d
)是两个分子例如蛋白质及其配体之间结合强度的量度。解离常数越小,两个分子结合得越紧密,两个分子之间的亲和力就越高。
[0039]
本文在eif4e活性的语境中所用的术语“抑制”是指与未受到抑制的eif4e的活性水平相比,eif4e的生物活性水平被破坏、降低或不存在。
[0040]
在肽或蛋白质的语境中,术语“大环(macrocycle)”和“大环(macrocyclic)”是指环状肽,其是含有键的环状序列的多肽链。这可以通过以下来实现:肽的氨基端和羧基端之间的连接,例如在环孢菌素中;氨基端和侧链之间的连接,例如在杆菌肽中;羧基端和侧链,例如在粘菌素中;或两条侧链或更复杂的排列,例如在鹅膏蕈碱中。
[0041]
本文所用术语“药物组合物”是指一种或多种本文所述化合物或其生理学/药学上可接受的盐或前药与其他化学成分,例如生理学/药学上可接受的载体和赋形剂的混合物。药物组合物的目的是促进将化合物施用于生物体。
[0042]
本文所用术语“药学上可接受的载体”是指本领域普遍接受的用于向哺乳动物例如人类递送生物活性剂(即所公开的单独或与说明书语境中任何公开化合物组合的抑制剂)的介质。这类载体通常根据本领域普通技术人员权限内决定和考虑的许多因素来配制。这些因素包括但不限于要被配制的活性剂的类型和性质;将给予含药剂组合物的个体;组合物的预期给药途径;以及所针对的治疗适应症。药学上可接受的载体包括水性和非水性液体介质,以及各种固体和半固体剂型。在活性剂之外,此类载体可包括许多不同的成分和添加剂,在制剂中包括这类额外成分出于多种原因,例如本领域普通技术人员公知的活性剂的稳定。药学上可接受的载体的非限制性实例是透明质酸及其盐和微球(包括但不限于聚(d,l)
‑
丙交酯
‑
共
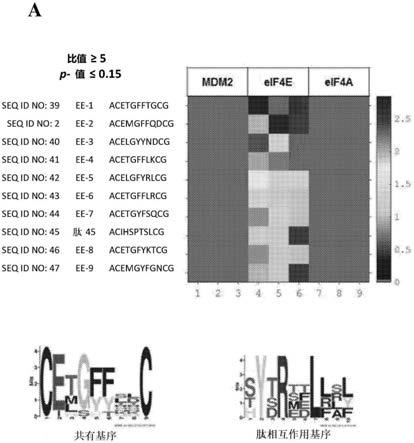
‑
乙醇酸共聚物(plga)、聚(l
‑
乳酸)(pla)、聚(己内酯)(pcl)和牛血清白蛋白(bsa))。本文所用术语“癌症”是指以以下为特征的许多疾病中的任何疾病:细胞不受控制的异常增殖,受影响细胞的局部扩散或通过血流和淋巴系统扩散到身体其他部位的能力(即转移),以及许多特征性结构和/或分子特征中的任何一个。癌症的实例包括但不限于乳腺癌、支气管癌、结肠癌、结肠直肠癌、肝癌、肺癌、前列腺癌、卵巢癌、脑癌、胰腺癌、头颈癌、胃癌和膀胱癌、非霍奇金淋巴瘤、白血病、神经母细胞瘤、黑素瘤、神经胶质瘤或胶质母细胞瘤。
[0043]
附图简要说明
[0044]
当结合非限制性实例和附图考虑时,参考详细描述会更好地理解本发明,其中:
[0045]
图1显示了下一代测序(ngs)噬菌体测序结果和荧光偏振竞争实验。使用c7c文库(可从neb购买)进行针对eif4e的ngs增强型噬菌体展示筛选。c7c=展示在m13噬菌体菌毛上的二硫化物约束环肽。图1a显示,鉴定了与eif4e相互作用的独特且新颖的基序,这与筛选线性12mer文库通常分离的肽相互作用基序形成对比。图1b显示了使用荧光偏振实验鉴定的结合位点。上图表明c7c肽不能置换含有肽相互作用基序的荧光标记肽。该基序存在于eif4e
‑
>4e
‑
bps(1、2和3)和eif4g1的蛋白质相互作用伴侣中。下图表明c7c大环ee
‑
02可以置换荧光标记的m7gtp。
[0046]
图2显示了色氨酸淬灭实验。该图展示了m7gtp和ee
‑
02如何具有非常不同的特征,这突出显示它们可以以不同的结合构象结合。phagsol是噬菌体衍生肽,经鉴定可与eif4e上的eif4g结合位点相互作用——含有规范的肽相互作用基序。
[0047]
图3显示了eif4e帽结合位点在有或没有m7gdp的复合物中的结构。图3a显示了m7gdp结合的eif4e结构(左)与未结合的结构(右)的比较。当结合m7gdp时,w56和w102的吲哚侧链插入核苷酸的鸟嘌呤环。eif4e的e103形成2个识别鸟嘌呤环的氢键。此外,存在于鸟嘌呤环n7上的甲基诱导季胺的形成,从而在环系统上赋予离域正电荷。这导致与w56和w102形成阳离子
‑
π相互作用,进一步稳定了结合构象。m7gdp的二磷酸盐尾被识别并与r157、
k159和k162形成静电相互作用。残基r112经由几个结构化水分子进一步识别磷酸盐部分。这种构象被称为“封闭”的构象。当eif4e未结合时,负责结合n7甲基取代的鸟嘌呤部分的区域周围会发生明显的构象变化。这些变化是w102的旋转、含w56的环的移动和e103旋转出帽结合位点。这种构象被称为“开放”状态。图3b显示了eif4e上规范肽和帽结合位点的位置。图3c显示了m7gdp结合的eif4e和“apo”eif4e结构的表面表示,证明了两种状态之间帽结合位点的形状和大小上的明显结构差异。
[0048]
图4显示了与ee
‑
02或m7gdp结合的eif4e结构的比较。图4a显示了ee
‑
02结合的eif4e的表面表示,表明其帽结合位点的形状和大小与“apo”eif4e的相似,并突出了与m7gdp结合的eif4e的帽位点的差异。图4b显示了ee
‑
02结合的eif4e结构与m7gdp结合的结构的比较。ee
‑
02结合eif4e的构象与eif4e的“apo”结构非常相似。ee
‑
02的e3直接与r112相互作用,m4与eif4e的s92在帽结合位点背面形成氢键,而残基f6和f7形成疏水相互作用。负责识别鸟嘌呤环系统的残基w56和w102从帽结合位点旋转出来。与m7gdp形成h键的e103也摆出(swung out)了帽结合位点,并且不参与ee
‑
02与eif4e的相互作用。此外,与m7gdp相反,ee
‑
02不直接与形成二磷酸结合口袋的残基(r157、k159和k162)相互作用。
[0049]
图5显示了晶体结构中由ngs噬菌体测序鉴定的相互作用基序的保守残基产生的关键相互作用。还突出显示了ee
‑
02与eif4e及其自身形成的关键氢键。
[0050]
上图中的所有图像均基于与eif4e复合的ee
‑
02的真实xtal
tm
结构。
[0051]
发明详述
[0052]
在第一方面,本发明涉及包含氨基酸序列cex1gx2x3x4x5c(seq id no:1)的肽,其中x1是选自由苏氨酸(t)、甲硫氨酸(m)或亮氨酸(l)组成的组的氨基酸;其中x2是选自由疏水性氨基酸、芳香族氨基酸、经修饰的疏水性氨基酸和经修饰的芳香族氨基酸组成的组的氨基酸;其中x3是选自由疏水性氨基酸、芳香族氨基酸、经修饰的疏水性氨基酸和经修饰的芳香族氨基酸组成的组的氨基酸;其中x4是任何氨基酸;其中x5是任何氨基酸;其中两个半胱氨酸残基通过二硫键连接;并且其中肽与延伸起始因子4e(eif4e)结合。
[0053]
在一个实施方案中,肽包含氨基酸序列cex1gffx4x5c(seq id no:13)。
[0054]
可以通过本领域公知的方法从细胞或反应容器中分离本发明的肽。也可以通过本领域公知的方法纯化本发明的肽。
[0055]
在一些实施方案中,可以化学合成本发明的肽,然后分离或纯化。例如,可以通过芴基甲氧羰基保护基策略(fmoc)固相化学合成来合成本发明的肽。接着进行由碘介导的氧化以形成约束肽的二硫键。
[0056]
在一个实施方案中,肽在mrna 5’帽结合位点与eif4e结合。该肽以高亲和力与eif4e的mrna 5'帽结合位点结合。在一个实施方案中,肽以小于约250nm、小于约200nm、小于约150nm、小于约100nm、小于约50nm的解离常数(k
d
)与mrna 5'帽结合位点结合。在一个实施方案中,肽在mrna 5’帽结合位点以小于100nm的k
d
与eif4e结合。
[0057]
肽可以通过不同于帽类似物(m7gdp)与5'帽结合位点相互作用的机制或模式与5'帽结合位点相互作用。
[0058]
在一个实施方案中,肽以不同于m7gdp的构象与eif4e帽结合位点结合。在另一个实施方案中,肽与eif4e的结合破坏了帽类似物与帽结合位点的结合。这转而抑制了eif4e的活性。
[0059]
在一个实施方案中,肽以开放构象与eif4e结合。“开放构象”是指,当第102和56位(w102和w56)的苯丙氨酸不与m7gdp相互作用并摆出结合位点时,第102和56位(w102和w56)处苯丙氨酸的构象。相比之下,“闭合构象”是指当w102和w56摆回(swung back)进入帽结合位点且m7gdp夹在它们之间时w102和w56的构象。
[0060]
在一个实施方案中,肽与eif4e的结合抑制eif4e活性。
[0061]
通常应理解,两个半胱氨酸残基之间的二硫键使肽环化。
[0062]
应当理解,本发明的环化肽与eif4e相互作用,并且当二硫键减少时,相互作用消除。本发明的肽代表负责与eif4e的这种相互作用的关键基序和约束肽的必要性。
[0063]
在一个实施方案中,本发明的包含氨基酸序列cex1gx2x3x4x5c(seq id no:1)的肽可以在x1位是甲硫氨酸(m)。在另一个实施方案中,x2和x3独立地选自由酪氨酸(y)、苯丙氨酸(f)或经修饰的苯丙氨酸组成的组。
[0064]
经修饰的苯丙氨酸是指芳香族环上的一个或多个基团被取代的苯丙氨酸。
[0065]
在另一个实施方案中,x4和x5独立地为谷氨酰胺(q)、天冬氨酸(d)、丙氨酸(a)、赖氨酸(k)、甘氨酸(g)或亮氨酸(l)。
[0066]
在又一个实施方案中,x5是天冬氨酸(d)。在又一个实施方案中,x5是d
‑
天冬氨酸或l
‑
天冬氨酸。
[0067]
在一些实施方案中,肽可以在n
‑
末端和/或c
‑
末端包含一个或多个另外的氨基酸残基。这些一个或多个另外的氨基酸残基可以来自在其上展示肽的噬菌体或噬菌体的一部分。在一个实施方案中,肽在n
‑
末端包含另外的丙氨酸残基和在c
‑
末端包含另外的甘氨酸残基。丙氨酸和甘氨酸残基是噬菌体的非随机部分,其存在于组成噬菌体文库的随机部分之外。
[0068]
在本发明的一个实施方案中,肽包含选自由acemgffqdcg(seq id no:2)、cemgffqdcg(seq id no:3)、acemgffadcg(seq id no:4)、acemgffkdcg(seq id no:5)、acemgffldcg(seq id no:6)、cemgffadc(seq id no:7)、acemgyfqdcg(seq id no:28)和acemgfyqdcg(seq id no:32)组成的组的氨基酸序列。
[0069]
在另一个实施方案中,肽由选自由以下组成的组的氨基酸序列组成:acemgffqdcg(seq id no:2)、cemgffqdcg(seq id no:3)、acemgffadcg(seq id no:4)、acemgffkdcg(seq id no:5)、acemgffldcg(seq id no:6)、cemgffadc(seq id no:7)、acemgyfqdcg(seq id no:28)和acemgfyqdcg(seq id no:32)。
[0070]
在另一个实施方案中,肽由氨基酸序列cemgffadc(seq id no:7)组成。
[0071]
本发明的肽可以与一种或多种另外的化合物缀合。在一个实施方案中,肽与一种或多种另外的肽缀合。
[0072]
在一些实施方案中,一种或多种另外的肽是细胞穿透肽。细胞穿透肽(cpp)是促进肽的细胞摄取的肽。cpp可分为合成的、嵌合的或蛋白质衍生的,或者可分为阳离子的、疏水的或两亲的。细胞穿透肽的实例包括但不限于聚精氨酸、反式激活转录激活剂(tat)、转运蛋白(transportan)和穿透蛋白(penetratin)。
[0073]
本发明还涉及本文所述的肽作为药物的用途。
[0074]
在一个实施方案中,本文所述的肽用于治疗与帽依赖性翻译失调相关的疾病状况。
[0075]
本发明还提供了包含本文所述肽的药物组合物。
[0076]
本发明还提供了本文所述的肽或药物组合物在制造用于治疗与帽依赖性翻译失调相关的疾病状况的药物中的用途。
[0077]
在一个实施方案中,帽依赖性翻译失调是异常eif4e表达或异常eif4e活性的结果。
[0078]
在一些实施方案中,异常eif4e表达是eif4e过表达。在其他实施方案中,异常eif4e活性是eif4e活性增加到生理水平以上。通常应当理解,eif4e的过表达意味着与正常生理表达水平相比,eif4e的表达增加。通常还应当理解,eif4e活性的增加可以通过4e
‑
bp1的磷酸化增加来介导。
[0079]
在一个实施方案中,与eif4e过表达相关的疾病状况是癌症。
[0080]
在另一个实施方案中,与帽依赖性翻译失调相关的疾病状况是强行控制(hijack)帽依赖性翻译的传染病。此类疾病的实例包括但不限于流感以及由冠状病毒引起的病毒性疾病。
[0081]
在又一个实施方案中,与帽依赖性翻译失调相关的疾病状况是脆性x染色体综合征或孤独症。
[0082]
本发明还提供了包含编码本文所述肽的核酸序列的载体。本发明还提供了用如本文所述载体转染的宿主细胞,其中所述宿主细胞表达如本文所述的肽。
[0083]
在缺乏本文没有具体公开的任何一个或多个要素、一个限制或多个限制的情况下,可以适当地实施本文说明性描述的本发明。因此,例如,术语“包含(comprising)”、“包括(including)”、“含有(containing)”等应被广泛而非限制地解读。此外,本文使用的术语和表达已被用作描述性而非限制性的术语,并且在使用此类术语和表达时无意排除所示和所述特征或其部分的任何等同物,但是应当认识到,在要求保护的本发明的范围内可以进行各种修改。因此,应当理解,虽然已经通过优选实施方案和任选的特征具体公开了本发明,但是本领域技术人员可以诉诸本文所公开的的其中体现的本发明的修改和变化,并且这类修改和变化视为在本发明的范围内。
[0084]
本文已经广泛地和一般性地描述了本发明。落入一般公开内容的每个较窄的类别和亚属分组也构成本发明的一部分。这包括本发明的一般性描述,该一般性描述带有从其中移除任何主题的附带条件或否定限制,无论本文是否具体引用了移除的材料。
[0085]
其他实施方案在所附权利要求和以下非限制性实施例内。此外,在根据马库什组描述本发明的特征或方面的情况下,本领域技术人员应当认识到,也因此根据马库什组的任何个体成员或成员子组描述了本发明。
[0086]
实验部分
[0087]
通过参考具体实施例进一步更详细地描述本发明的非限制性实施例和对比例,不应将具体实施例解释为以任何方式限制本发明的范围。
[0088]
方法
[0089]
噬菌体展示法和荧光竞争实验
[0090]
荧光各向异性测定和k
d
测定
[0091]
一般的测定设计
[0092]
针对50nm羧基荧光素(fam)标记的示踪肽(kkrysrdfllalqk
‑
(fam))(seq id no:
8)或50nm荧光素标记的m7gtp
‑6‑
fam滴定纯化的eif4e。示踪肽和fam标记的m7gtp的k
d
(解离常数)通过使用以下方程将它们各自的实验滴定拟合到1:1结合模型来确定:
[0093][0094]
其中[p]是蛋白质浓度,[l]是标记肽浓度,r是测量的各向异性,r0是游离肽的各向异性,r
b
是eif4e
‑
示踪肽复合物的各向异性,[l]
t
是fam标记肽的总浓度,[p]
t
是eif4e总浓度。针对示踪肽与eif4e相互作用测定的测定k
d
为50.3nm,而m7gtp
‑6‑
fam的k
d
为149.0nm。两者稍后都用于在竞争实验中随后测定k
d
。
[0095]
然后通过竞争性各向异性实验测定各种分子的表观k
d
值。
[0096]
4e:4g位点处的结合评估
[0097]
进行滴定,其中eif4e的浓度恒定在200nm,标记肽的浓度恒定在50nm。然后针对fam标记的肽和eif4e的复合物滴定竞争分子。通过将实验数据拟合到下面所示的方程来确定表观k
d
值:
[0098][0099]
d=k
d1
+k
d2
+[l]
st
+[l]
t
‑
[p]
t
[0100]
e=([l]
t
‑
[p]
t
)k
d1
+([l]
st
‑
[p]
t
)k
d2
+k
d1
k
d 2
[0101]
f=
‑
k
d1
k
d2
[p]
t
[0102][0103]
[l]
st
和[l]
t
分别表示标记配体和总未标记配体输入浓度。k
d2
是未标记配体与蛋白质之间相互作用的解离常数。在所有类型的竞争实验中,假设[p]
t
>[l]
st
,否则会始终存在大量游离的标记配体并会干扰测量。k
d1
是各自实验所用的标记肽的表观k
d
。将示踪肽溶解于1mm的dmso中,并稀释到实验缓冲液中。使用envision multi
‑
label reader(perkinelmer)进行读数。实验在pbs(2.7mm kcl,137mm nacl,10mm na2hpo4和2mm kh2po4(ph 7.4)),3%dmso(v/v)和0.1%tween 20缓冲液中进行。所有滴定一式三份进行。使用prism 4.0(graphpad)进行曲线拟合。
[0104]
帽类似物结合评估
[0105]
进行滴定,其中eif4e的浓度恒定在250nm,标记的m7gtp的浓度恒定在50nm。然后针对fam标记的m7gtp和eif4e的复合物滴定竞争分子。通过将实验数据拟合到下面所示的方程来确定表观k
d
值:
[0106][0107]
d=k
d1
+k
d2
+[l]
st
+[l]
t
‑
[p]
t
[0108]
e=([l]
t
‑
[p]
t
)k
d1
+([l]
st
‑
[p]
t
)k
d2
+k
d1
k
d 2
[0109]
f=
‑
k
d1
k
d2
[p]
t
[0110][0111]
[l]st和[l]t分别表示标记配体和总未标记配体输入浓度。k
d2
是未标记配体与蛋白质之间相互作用的解离常数。在所有类型的竞争实验中,假设[p]
t
>[l]
st
,否则会始终存在大量游离的标记配体并会干扰测量。k
d1
是各自实验中所用的标记肽的表观k
d
。将fam标记的m7gtp以1mm提供于tris
‑
hcl ph7.5(jena biosciences)中,并稀释到实验缓冲液中。使用envision multi
‑
label reader(perkinelmer)进行读数。在pbs(2.7mm kcl、137mm nacl、10mm na2hpo4和2mm kh2po4(ph 7.4))、3%dmso(v/v)和0.1%tween 20缓冲液中进行实验。所有滴定一式三份进行。使用prism 4.0(graphpad)进行曲线拟合。
[0112]
ngs增强型噬菌体展示
[0113]
针对eif4e选择c7c二硫化物约束的噬菌体文库
[0114]
噬菌体库
[0115]
c7c噬菌体文库购自new england biolabs。该文库是具有二硫化物约束环的随机肽组合文库,该环与m13噬菌体的次要衣壳蛋白(piii)融合。展示的肽以ac
‑
x7
‑
cgggs(x=随机氨基酸)(seq id no:9)的形式在piii的n末端表达。两个半胱氨酸在氧化条件下自发形成二硫桥以约束肽序列。该文库由大约1x 109个电穿孔(即独特的)序列组成。
[0116]
eif4e固定和噬菌体选择
[0117]
将96孔聚苯乙烯板用链霉亲和素溶液包被(每孔150μl,20μg/ml,并在4℃孵育过夜。3.然后去除涂层溶液。然后将96孔板上的孔用200μl结合缓冲液(50mm tris
‑
hcl、150mm nacl(ph 7.4))洗涤4次。洗涤后,向孔加载150μl生物素标记的蛋白(在结合缓冲液中5
‑
10mg/ml)。链霉亲和素对照孔仅加载150μl结合缓冲液。然后将96孔板在室温孵育15min,然后倒出其中的内容物。然后用200μl结合缓冲液将孔洗涤4次。洗涤后,用200μl封闭缓冲液((50mm tris
‑
hcl,150mm nacl(ph 7.4).0.01%bsa(w/v),0.1%tween20(v/v))在室温将孔封闭30min。同时,在封闭缓冲液中以1x 10
12
个pfu/ml制备噬菌体文库,并且还将其在室温孵育30min。然后从96孔板中倒出封闭缓冲液,并将100μl噬菌体文库分布在各孔之间,每孔约10
11
个pfu。然后将板在室温孵育1小时。然后通过从各个孔中用移液管吸取去除未结合的噬菌体。然后用200μl洗涤缓冲液将孔洗涤8次。然后将结合的噬菌体与100μl洗脱缓冲液(0.2m甘氨酸
‑
hcl,ph 2.2+0.1%(w/v)bsa)在室温孵育9min。然后将含有洗脱噬菌体的上清液单独转移到含有15μl中和缓冲液(1m tris
‑
hcl,ph 9.1)的单独管中。
[0118]
噬菌体扩增和m13 dna分离
[0119]
将过夜培养的大肠杆菌(er 2738菌株)稀释100倍,并将3ml培养物分配到每个15ml培养管中。在37c将培养物孵育4
‑
4.5小时,同时以230rpm振荡。孵育后,将管在4℃以4000rpm离心15min以沉淀细菌细胞。为了沉淀噬菌体,将每个管中的上清液倒入含有mp缓冲液(30μl,100x)的单独的5ml falcon管中,并涡旋。将700μl沉淀的噬菌体溶液加载到单独的qiagen离心柱中,并以8000rpm离心30秒。丢弃由此产生的流通液(flow
‑
through)。在这一步之后,将700μl缓冲液pb施加到每个离心柱上,并以8000rpm离心30s。再次丢弃流通液并重复此步骤。完成此步骤后,向每个离心柱添加700μl缓冲液pe。将每个柱以8000rpm旋转离心30s,并丢弃收集管中的流通液。重复离心步骤,但将旋转离心延长至2min以去除任
何残留的pe缓冲液。将离心柱转移到新鲜的1.5ml eppendorf管中。将dna添加eb缓冲液(50μl)洗脱到每根柱子中,然后等待1min,接着以800rpm离心30s。有关缓冲液和程序的更多详细信息,请参阅m13qiaprep spin m13试剂盒手册(qiagen,ltd)。
[0120]
mp缓冲配方
[0121]
1.在室温(21℃)将3.3g一水柠檬酸溶解在3ml高纯水中
[0122]
2.通过以200rpm搅拌,将溶液孵育5min
[0123]
3.使用注射器用0.2μm无菌过滤器过滤溶液,得到最终体积为6ml的buffer mp。
[0124]
条形码和文库扩增
[0125]
15个带条形码的反向引物设计有与illumina测序兼容的衔接子。用位于可变区侧翼的带条形码的引物对文库dna进行pcr扩增。ssm13噬菌体dna的每个洗脱液都是唯一编码的。简而言之,使用缓冲液、200μm每种dntp、1.5mm mgcl2、0.5μm正向引物、0.5μm带条形码的反向引物和一个单位的高保真dna聚合酶,总体积为25μl,扩增文库dna(15个样品,每个50ng)。使用以下热循环仪程序进行pcr:95℃30s,然后是95℃10s、60.5℃15s和72℃30s,25个循环,然后在72℃进行5min最终延伸,然后保持在4℃。使用低分子量dna ladder作为标准(neb,#n3233s),通过在tris
‑
硼酸盐
‑
edta缓冲液中以2%(w/v)琼脂糖凝胶在100伏运行约45min,对来自pcr的dsdna片段进行定量。将dsdna片段(15个样品,每个样品40ng)合并在一起,并在sizeselecttm 2%琼脂糖凝胶(invitrogen,#g6610
‑
02)上纯化。参考ladder,用不含rna酶的水收集对应于121bp的所需条带。根据制造商的说明,使用picogeen(thermofisher)测定经纯化的dna的dna浓度。
[0126]
dna模板制备和illumina测序(nextseq)
[0127]
测序由axil scientific进行。将合并的dna与illumina芯片杂交,进行桥式扩增,然后使用illumina技术测序。500mid output kit(150个循环)(目录号fc
‑
404
‑
1001)和500/550mid output kit v2(150个循环)(目录号fc
‑
404
‑
2001)都被用于在正向(f)和反向(r)方向对读数进行测序。使用内部开发的python脚本,使用双端分析处理fastq文件。
[0128]
差异富集和序列标识生成
[0129]
分析和解析fastq文件以通过相关条形码将序列分离到它们各自的选择中。然后确定可变域编码的每个独特肽序列的频率,并通过选择中的序列总数归一化。对于每个重复,例如eif4e重复1
‑
3和链霉亲和素对照1
‑
3,重复该过程。在特定重复中未观察到的序列被指定为归一化频率为零。通过将特定序列的平均归一化频率(来自针对该选择的重复集)除以对照重复集中的频率,来确定每个序列的富集比率。由于在取比值时分母不能为零,因此在计算比值之前,未在所有三个重复中出现的序列被赋予任意值(例如0.0001)。使用不等方差斯氏t检验(welch检验)评估比率的显著性(p值)。然后为p值和富集比率的任意确定值生成热图。使用的典型值为p值<0.1和比率>5.0。序列按其在热图中的富集值排序。热图确定了从特定选择中分离出的序列,相对于从对照选择(或非相关选择组)中分离出的序列,这些序列的丰度显著增加。使用weblogo(https://weblogo.berkeley.edu/)从富集的序列中生成序列标识。
[0130]
色氨酸淬灭实验
[0131]
使用多板读数器和黑色哑光96孔板进行色氨酸淬灭实验。用1μm纯化的eif4e针对以下化合物的一系列浓度制备样品(100μl):m7gtp、phagesol(kkrysrdqlval)(seq id no:10)或ee
‑
02。测试的浓度范围为0.01
‑
20μm。用290nmuv光激发样品,以最大限度地减少酪氨酸荧光的影响。在355nm的发射波长下测量色氨酸。
[0132]
eif4e:ee
‑
02的结晶
[0133]
结晶
[0134]
使用坐滴法通过蒸汽扩散使eif4e:ee
‑
02复合物结晶。结晶滴含有浓度分别为150μm和300μm的eif4e和ee
‑
02。在48孔intelli
‑
plate(汉普顿研究)中设置坐滴,并将1μl蛋白质样品与1μl母孔溶液混合。晶体在0.2m硫酸铵0.1m tris醋酸三钠ph 4.6和25%peg 4000中生长一周。为了在100k收集x射线数据,将晶体转移到含有20%(v/v)甘油的等效母液(mother liquor solution)中,然后在液氮中快速冷冻。
[0135]
数据收集和细化
[0136]
使用ccd探测器,在光束线mx1的澳大利亚同步加速器上收集数据。晶体衍射到的分辨率,并使用xds求积分和缩放。使用去除水和其他结合成分的人类eif4e结构(pdb登录代码:4bea)作为搜索模型,通过分子置换和程序phaser来解析eif4e的二元复合晶体的初始相。使起始模型接受刚体细化,然后在coot中进行手动模型构建的迭代循环,并在refmac 6.0中进行限制细化。
[0137]
一般方法
[0138]
eif4e的表达、重折叠和纯化(用于荧光偏振(fp)、表面等离子体共振(spr)和晶体学实验)
[0139]
用含有全长eif4e克隆的pet11d表达质粒转化rossetta plyss感受态细菌。两种材料均由dundee的cyclacel ltd.提供。使表达全长eif4e构建体的细胞在37℃的lb培养基中生长至od
600
约为0.6,并用1mm iptg开始诱导eif4e。立即将培养物置于振荡培养箱(shaker
‑
incubator)中,在37℃持续3h。通过离心收获细胞,并将细胞团重悬于50mm tris ph 8.0、10%蔗糖中,然后进行超声处理。
[0140]
将超声处理的样品在4℃以17,000g离心10min。将所得沉淀重悬于tris/triton缓冲液(50mm tris ph 8.0、2mm edta、100mm nacl、0.5%triton x
‑
100)中。然后将样品在4℃以25,000g离心15min,并将沉淀重悬于tris/triton缓冲液中。重新离心后,将剩余的沉淀溶解在6m盐酸胍、50mm hepes
‑
koh ph 7.6、5mm dtt中。然后将样品的蛋白质浓度调整为1mg/ml。
[0141]
通过1/10稀释将变性蛋白质重折叠到由20mm hepes
‑
koh、100mm kcl和1mm dtt组成的重折叠缓冲液中。使用amersham pd10柱将重折叠的蛋白质浓缩并脱盐到重折叠缓冲液中。将eif4e蛋白质样品在monoq柱上运行,并用1m kcl梯度洗脱。eif4e在约0.3m kcl处洗脱为尖峰。
[0142]
表面等离子体共振(用于结构活性关系(sar)表)
[0143]
对于肽储备溶液,将化合物溶解在100%dmso中至浓度为10mm;在分析之前,立即将肽储备溶液进一步稀释到dmso和/或运行缓冲液中。运行缓冲液由10mm hepes ph 7.6、0.15m nacl和0.1%tween20组成。将肽储备溶液/dmso稀释的肽溶液稀释到1.03
×
运行缓冲液中,以制备dmso终浓度为3%的肽溶液。通过将样品进一步稀释到含有3%dmso的运行
缓冲液中,达到肽的工作浓度。在biacore t100机器上进行表面等离子体共振实验。
[0144]
将纯eif4e固定在cm5传感器芯片上。通过注射100mm hcl 6s调节cm5芯片,然后注射0.1%sds 6s,并通过以100μl/分钟的流速注射50mm naoh 6s来结束。使用nhs(115mg ml
‑1)和edc(750mg ml
‑1)的混合物以10μl min
‑1激活传感器芯片表面,持续7min。用10mm醋酸钠缓冲液(ph 5.0)将纯化的eif4e稀释至终浓度为0.5μm,并且m7gtp以2:1的比例存在,以稳定eif4e。固定在激活表面上的eif4e的量通过改变蛋白质溶液的接触时间来控制,约为1000ru。固定蛋白质后,利用注射1m乙醇胺(ph 8.5)7min(10μl min
‑1)来淬灭过量的活性琥珀酰亚胺酯基团。
[0145]
首先注射六个缓冲空白以完全平衡仪器,然后进行溶剂校正曲线,随后进一步进行2个空白注射。通过将不同量的100%dmso添加到1.03x运行缓冲液以生成一系列dmso溶液(分别为3.8%、3.6%、3.4%、3.2%、3%、2.85%、2.7%和2.5%)来设置溶剂校正曲线。使用50μl/min的流速将化合物注射60s,并监测解离180s。数据收集率为10hz。k
ds
使用biaevaluation软件(biacore)来确定,并根据平衡时eif4e包被的cm5芯片的响应以及根据每个肽的解离和结合相数据的动力学来计算。将平衡和动力学数据均拟合为1:1结合模型。由三个单独的滴定确定每个个体肽的k
d
。在每次滴定中,至少复制两个浓度点以确保芯片表面的稳定性和坚固性。
[0146]
用于生物素化的eif4e的gst亲和纯化
[0147]
通过bamh1和nde1双消化将全长eif4e连接到gst融合表达载体pgex
‑
4(ge lifesciences)中。然后用带有gst标签的eif4e融合构建体转化bl21 de3感受态细菌。使表达gst融合构建体的细胞在37℃的lb培养基中生长至od
600
为约0.6,并在室温下以1mm进行诱导。通过离心收获细胞,将细胞团重悬于50mm tris ph 8.0、10%蔗糖中,然后进行超声处理。将超声处理的样品在4℃以17,000g离心60min。将上清液施加到在具有1mm dtt的洗涤缓冲液(磷酸盐缓冲盐水,2.7mm kcl和137mm nacl,ph 7.4)中预平衡的5ml ff gst柱(amersham)上。然后用6体积的洗涤缓冲液进一步洗涤柱子。然后通过用precission(ge lifesciences)蛋白酶切割从柱中纯化eif4e。将50个单位的凝血酶蛋白酶(在具有1mm dtt缓冲液的一柱体积pbs中)注射到柱上。使切割反应在4℃进行过夜。然后用洗涤缓冲液从柱子上洗脱切割的蛋白质。使用sds
‑
page凝胶分析蛋白质级分并使用millipore的centricon(10kda mwco)浓缩器进行浓缩。然后将eif4e蛋白质样品透析到含有20mm hepes
‑
koh、100mm kcl和1mm dtt的缓冲溶液中。然后使eif4e蛋白质样品通过monoq柱并用1m kcl梯度洗脱。eif4e在约0.3m kcl处洗脱为尖峰。使用uv吸光度测定蛋白质浓度。
[0148]
用于固定和(n)噬菌体展示的eif4e的分选酶介导的生物素化
[0149]
使用分选酶介导的连接将生物素标签掺入到gs
‑
eif4e上。在连接缓冲液(50mm tris、1500mm nacl、10mm tcep(ph 8.0))中,以1000μl的体积混合以下试剂:40μm gs
‑
eif4e、5μm srta61
‑
206
‑
his6(8m)、200μm生物素
‑
kggglpet
‑
gg
‑
ohse(ac)
‑
酰胺。将样品在室温下孵育4小时。通过sds
‑
page凝胶分析2μl样品的标记效率。分析后,然后纯化反应样品以除去除eif4e之外的所有反应组分。样品用50μl预洗涤的his
‑
tag磁珠(qiagen)重悬浮,并在室温在滚筒上孵育5min。7.将管子在磁铁上放置2min,然后将含有生物素化蛋白的上清液转移到新鲜的eppendorf管中。使用slide
‑
a
‑
lyzer盒(10k mwco)对2l pbs透析生物素化蛋白质(4小时),然后将缓冲液改为50mm tris、150mm nacl(ph7.4),并透析过夜。测量蛋
白质浓度。等分并快速冷冻蛋白质样品。
[0150]
结果
[0151]
在帽结合位点结合eif4e的新型大环肽
[0152]
针对n末端生物素化的eif4e淘选噬菌体展示文库。所用的噬菌体展示文库由受两个半胱氨酸残基(acxxxxxxxc
–
gggs)(seq id no:9)之间的二硫键约束的7个随机氨基酸组成,其通过ggs接头与m13噬菌体的次要衣壳蛋白(piii)融合。该文库是从new england biolabs(neb)商业采购的。eif4e通过肽连接生物素化,由此使用分选酶将生物素标记肽(生物素
‑
lpetgg)(seq id no:37)与eif4e上的n末端甘氨酸融合。通过使用gst标签进行纯化和使用凝血酶去除gst标签,产生带有n末端甘氨酸eif4e。将生物素化的eif4e固定在链霉亲和素包被的板上。
[0153]
通过针对eif4e进行3x选择和3x对照选择(针对mdm2和eif4a二者),进行下一代测序(ngs)增强型噬菌体展示。如方法中所述进行选择。使用illumina nextseq技术(参见方法)对噬菌体进行分离、扩增和测序。使用内部python脚本分析数据并鉴定了一组具有新的eif4e相互作用基序的独特肽(图1)。
[0154]
鉴定的共有基序由c
‑
e
‑
(t/m/l)
‑
g
‑
f
‑
f
‑
x
‑
x
‑
c(seq id no:11)组成。x表示任何氨基酸。使用由针对eif4e的随机12mer线性肽(neb)组成的噬菌体文库进行平行ngs增强噬菌体选择。然而,该库仅鉴定了在eif4e上的eif4g结合位点相互作用的eif4e良好相互作用结合基序。(yxrxxlφ,φ=任何疏水性氨基酸)(seq id no:12)。
[0155]
根据这些初始结果可以预测,新的肽基序可能在蛋白质上的eif4g结合位点、或帽结合位点或其他地方通过不同的结合姿势与eif4e相互作用。通过使用两种类型的竞争性荧光偏振实验,其中一种使用fam标记的帽类似物,另一种使用fam标记的eif4g1衍生肽,在eif4e上初始地描绘了结合位点(图1)。这些实验证实,ee
‑
02(acemgffqdcg)(seq id no:2)(图1)是所鉴定的组中最有效的肽,并证实这种新的相互作用基序破坏了帽类似物与eif4e的结合。
[0156]
ee_02的结构与帽类似物的结构明显不同,帽类似物由在鸟嘌呤环的n7位置具有甲基取代的鸟嘌呤三磷酸分子组成。这直接表明大环肽可以通过非常不同的机制与帽结合位点相互作用。
[0157]
为了证实这一假设,进行了色氨酸淬灭实验,其中针对eif4e滴定了ee
‑
02、m7gtp或eif4g1衍生肽(图2)。
[0158]
与其他滴定剂相比,phagesol肽产生的色氨酸淬灭量明显更低。这一结果是预料之中的,因为只有一个色氨酸残基(w73)位于eif4g结合位点附近,特别是与m7gtp/m7gdp相比,m7gtp/m7gdp在帽结合位点结合,并插入两个色氨酸残基(w56和w102)之间,因此导致明显更多的淬灭。然而,也在帽结合位点结合的ee
‑
02产生与m7gtp/m7gdp明显不同的特征。这种差异表明,ee
‑
02与eif4e的相互作用通过不同的模式发生,并且在它如何影响帽结合位点方面可能是变构的。
[0159]
为了解决这个问题,进行了结构研究,并成功解决了与eif4e结合的ee
‑
02的晶体结构(图3和4)。
[0160]
结构研究表明,ee
‑
02直接与帽结合位点相互作用,不会插入色氨酸环之间。该结果解释了在色氨酸猝灭实验中观察到的不同曲线。此外,ee
‑
02大环与eif4e结合,而不会模
仿m7gdp产生的磷酸盐尾或阳离子
‑
π相互作用(图3、4和5)。
[0161]
ee
‑
02通过4个不同的口袋与eif4e相互作用(图5)。大环本身形成β
‑
发夹转入帽结合位点,使残基e3、e4、f6和f7能够突入(projection)到图5所示的相应口袋中。
[0162]
还对ee
‑
02进行了丙氨酸扫描诱变,以确认在晶体结构中确定的关键相互作用(表2)。
[0163]
表2:ee
‑
02的丙氨酸扫描诱变
[0164][0165][0166]
使用方法中所述的表面等离子体共振(spr)确定k
d
。丙氨酸扫描证实残基e3、m4、g5、f6和f7对与eif4e结合都是关键的。g5对取代的敏感性突出了其在允许在结合口袋中形成具有正确几何形状的β
‑
发夹中的重要性。用a取代q8不影响结合。然而,用a置换d9导致结合减少两倍。
[0167]
表3:ee
‑
02的e3和m4位点上的靶向诱变
[0168][0169]
e4不能被d4置换,这强调了羧基正确取向以与eif4e的r112相互作用的重要性。m4被较短和较小的脂肪族基团(如i和l)替换导致结合丧失。m4也被替换为大小与甲硫氨酸相似的非天然脂肪族氨基酸正亮氨酸和环丁基丙氨酸。然而,针对这些分子确定的针对eif4e的k
d
仍然降低,这突出了硫原子与s92相互作用的重要性。
[0170]
表4:残基g5的靶向诱变
[0171][0172]
用不同的氨基酸(β
‑
丙氨酸(ba)、肌氨酸(sar)、p)置换g5,以探索围绕该位置的不同立体化学限制。所有这些都导致与eif4e结合的大环显著降低,进一步突出了g5在允许β发夹形成并将关键相互作用残基定向到其最佳位置以结合eif4e方面的作用。
[0173]
表5:ee
‑
02中f6和f7位的靶向诱变
[0174][0175]
通过探索f6和f7上的环取代基,可以逐步提高与eif4e的结合。用y6置换f6或用y7置换f7产生两个比ee
‑
02结合更紧密的分子(分别为ee
‑
30和ee
‑
35)。这些结果表明可以进一步探索这些位置以产生更好的结合类似物。
[0176]
表6:ee
‑
02中q8位的靶标诱变
[0177][0178]
ee
‑
02的第8位没有显示对特定残基的严格偏好。这与在晶体结构中与eif4e没有相互作用的事实相联系。然而,g突变的结合略有下降表明该位置的刚性化策略可以通过熵减少机制改善结合。
[0179]
表7:对ee
‑
02的截短研究
[0180][0181]
这些研究旨在发现可以结合eif4e的分子量(mw)最小的大环。此外,研究了大环的n端加帽,以鉴定乙酰化与游离胺对eif4e结合的影响。这导致ee
‑
44被鉴定,非文库噬菌体氨基酸位置a1和g11可以被去除,并且乙酰化显著降低了大环的结合。ee
‑
44由于其小mw和降低的尺寸,是用于合理设计具有改善的穿透性(permeability)的大环的更合适起点。
[0182]
表8:序列表总结
[0183]
[0184][0185]
等同
[0186]
上述实施例是为了说明本发明而提出的,不应被解释为对本发明的范围施加任何限制。很明显,在不脱离本发明的基本原理的情况下,可以对上述和实施例中说明的本发明的具体实施例进行许多修改和改变。所有此类修改和改变均旨在被本技术涵盖。
完整全部详细技术资料下载
当前第1页 1 2
相关技术
- 蚕虫草活性成分的制备工艺及其...
- 一种改性聚乙烯醇及其制备方法...
- 工程化CD25多肽及其用途的...
- 一种替考拉宁的脱色方法与流程
- 一种皮制品加工用表面压纹装置...
- 用于过继性T细胞疗法的中央记...
- 一种水性UV表涂处理耐刮PP...
- 一种皮革毛皮制品自动开料设备...
- 一种耐老化橡胶的制备方法与流...
- 用于RTV硅橡胶的聚倍半硅氧...
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1